接種時期選定の参考 (新)

親和性成熟とクラススイッチの説明には、コロナのワクチンを自分で考えるうえで少し役立つ内容が含まれています。
初めての人にとっては少し難しいので、先に「3-1-3.抗原提示」と「3-1-4.T細胞受容体」が終わってからここへ戻るほうがわかりやすいでしょう。
親和性 とは「抗体とエピトープ(抗原決定基)との結合力の強さ」を意味しています。
抗体とエピトープとの相性がよく、結合力が強いことを「親和性が高い」といいます。
男女関係と同じですね。
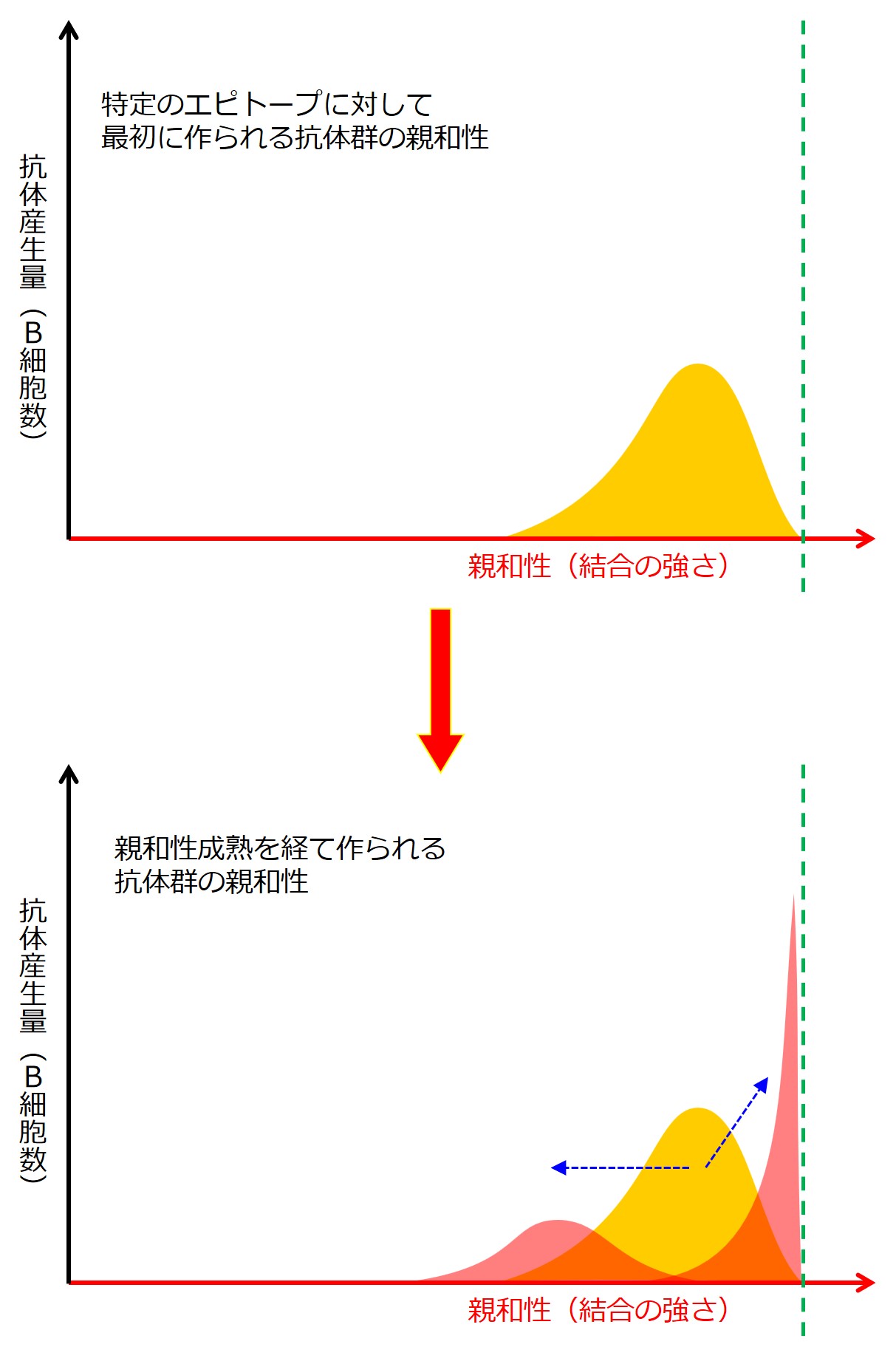
親和性成熟という言葉から、抗体が何度も作り直されて結合力が強くなっていく様子を思い浮かべられるかもしれませんが、・・・違います。
親和性成熟 は、エピトープに対する結合のバリエーションが拡がることと、狭まることの両方を意味しています。
エピトープに対する結合のバリエーションが拡がるということは、当該エピトープに対する結合力は弱まるが、他の似たような(つまり少し変異した)エピトープに結合できるようになることを意味し、エピトープに対する結合のバリエーションが狭まることは、当該エピトープに対する結合力がどんどん強くなり、当該エピトープにガッチリと結合するような抗体を産生できるようになることを意味しています。
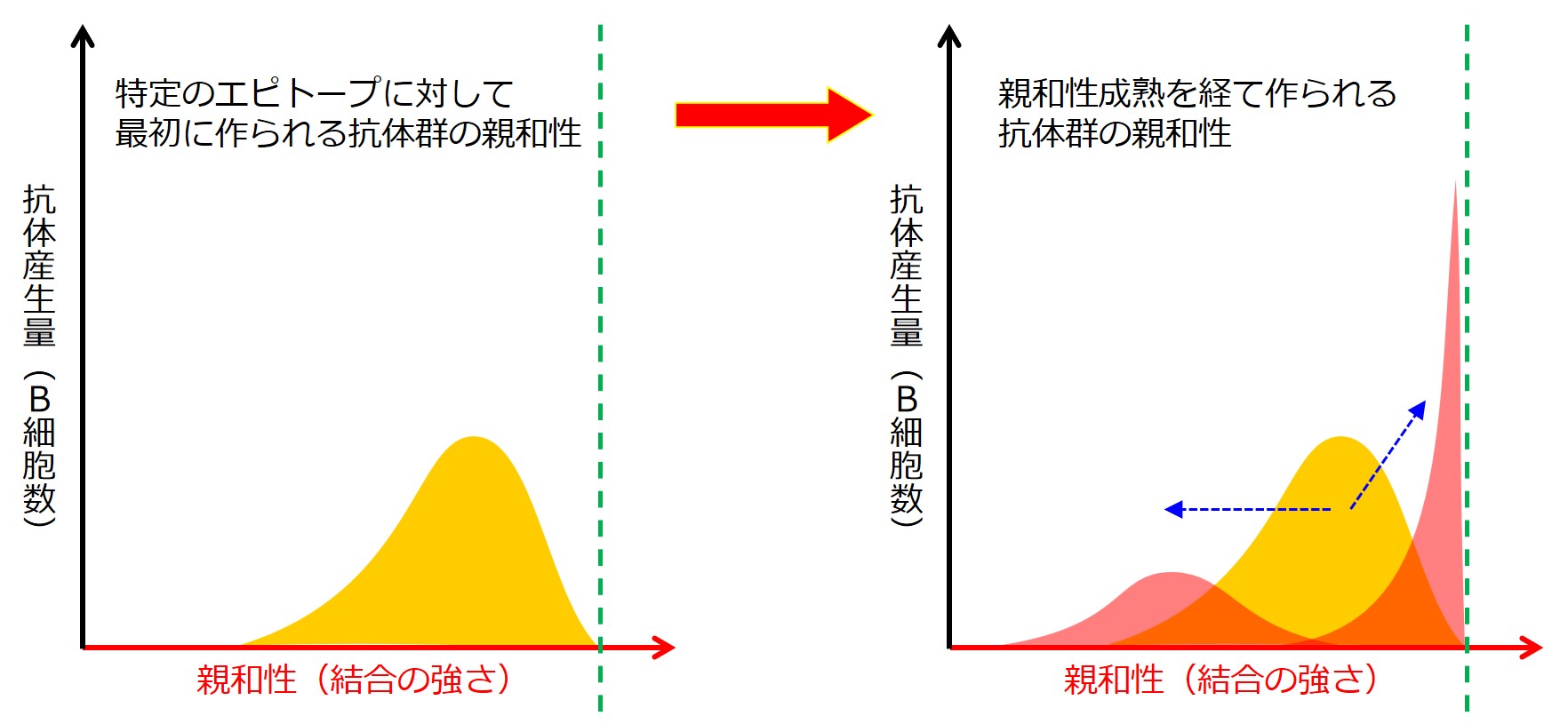
緑の破線は、このエピトープに最も強く結合する抗体(産生B細胞)の位置を示しています。
このグラフは、あとで描き直しますので、この段階であれこれ深読みしないようにしてください。
先にわかりやすいたとえを紹介します。
親和性成熟は子供たちの勉強と同じです。学校の教科書を何度も繰り返し読むことで、知識の広がりと深さが変化します。好きでない領域でも知識は増えていくし、また好きな部分はとても詳しくなるでしょう。
男女関係も同じですね。
見知らぬ数十人で会社に就職したら、同期みんなと仲良くなるでしょうが、もし相性の良い相手を見つけたら特に仲良くなっていくでしょう。親和性成熟は、仲良しの幅を拡げることと、仲良しの度合いを深めていくことの両方を意味しています。
クラススイッチ は、「3-1-2-3.抗体の種類」で述べたように、免疫グロブリン(Ig)のクラスが「M→G」とか、「M→A」とか、「G1→G4」などのように切り替わることです。
クラススイッチでは抗体重鎖の定常領域(CH)が入れ替わります。
では、抗体の親和性成熟とクラススイッチを説明します。
抗体産生を担うB細胞は、骨髄(Bone marrow)で生まれ、骨髄で教育を受けます。
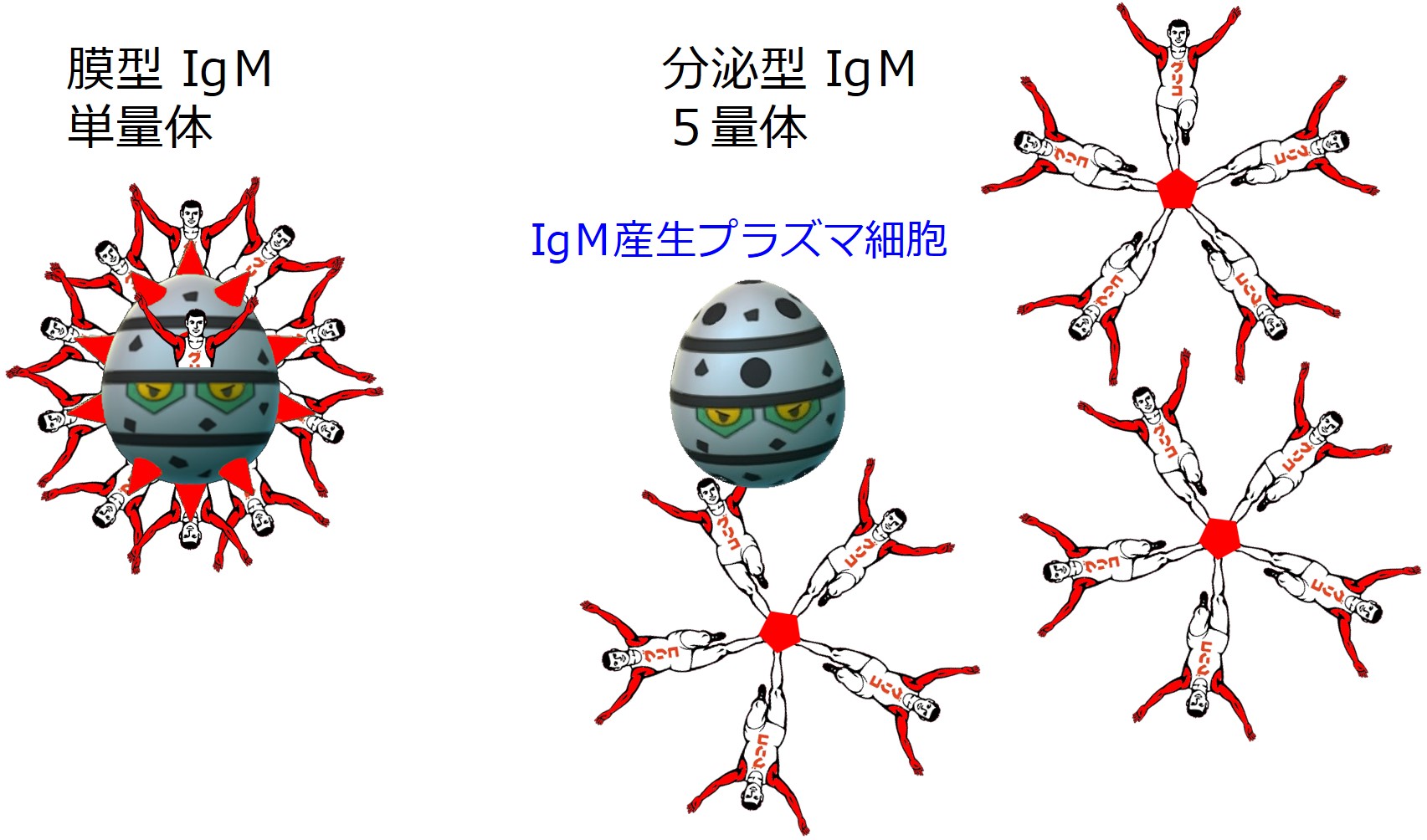
このときB細胞は特定のエピトープに対応した IgM を産生し、細胞膜表面に同じ IgM(単量体)をたくさん配置しています(膜型IgM)。
骨髄での教育課程を修了したけれども、まだ実際の抗原と出会って活性化していない幼若なB細胞を「ナイーブなB細胞」と呼びます。要するに童貞や処女ということです。
T細胞は、B細胞と同じように骨髄で生まれますが、心臓の前にある胸腺(Thymus)で教育を受けます。胸腺での教育を終えたけれど、まだ自分の担当する抗原と出会って活性化していない幼若なリンパ球を「ナイーブT細胞」と呼びます。
社交的でよくしゃべる子はナイーブヘルパーT細胞になり、寡黙だけど殺しのうまい子はナイーブキラーT細胞になります。
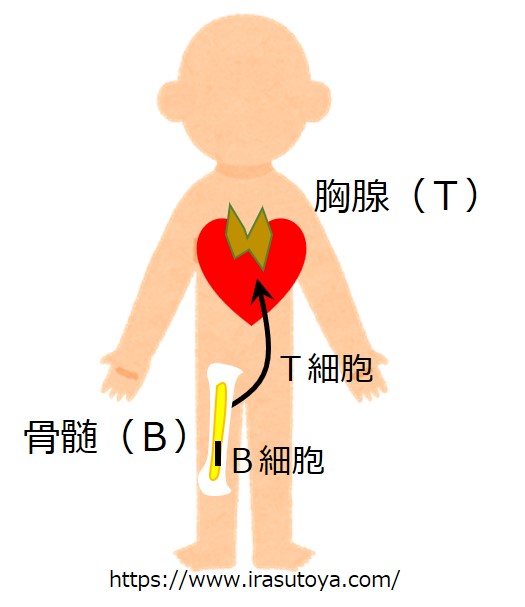
さて、骨髄を出たナイーブB細胞と胸腺を出たナイーブT細胞は、血液やリンパ液の流れに乗って、全身にあるいろいろなリンパ組織(リンパ節、脾臓など)を巡ります。
血液の液体成分は毛細血管の壁から常に染み出ており、リンパ液となります。リンパ液はリンパ管を通って体の中央へ向かい、途中のリンパ節に流れ込みます。
リンパ節には多数のリンパ球がたむろしています。
リンパ節の中は、ナイーブB細胞の多い区域(B細胞領域)とナイーブT細胞の多い区域(T細胞領域)とに分かれています。B領域は濾胞(ろほう:follicle)とも呼ばれます。
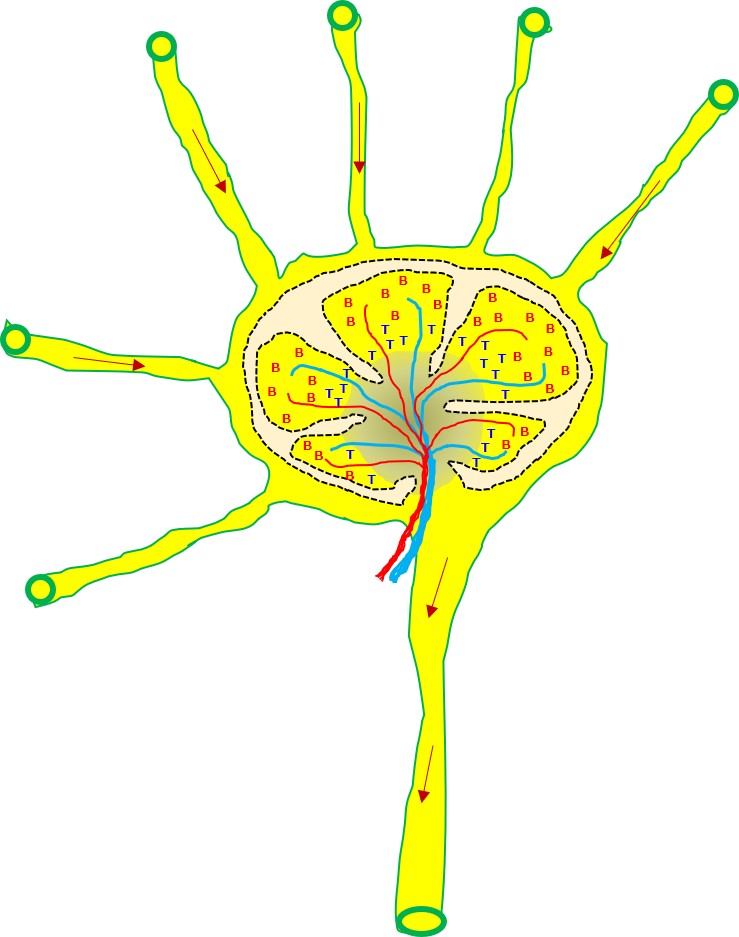
病原体が身体内に侵入すると、そこで食細胞の攻撃を受けます。病原体そのものや破壊された断片はリンパの流れに乗ってリンパ節に運ばれます。
抗原断片には1個 ~ 複数個の異なるエピトープがあることでしょう。
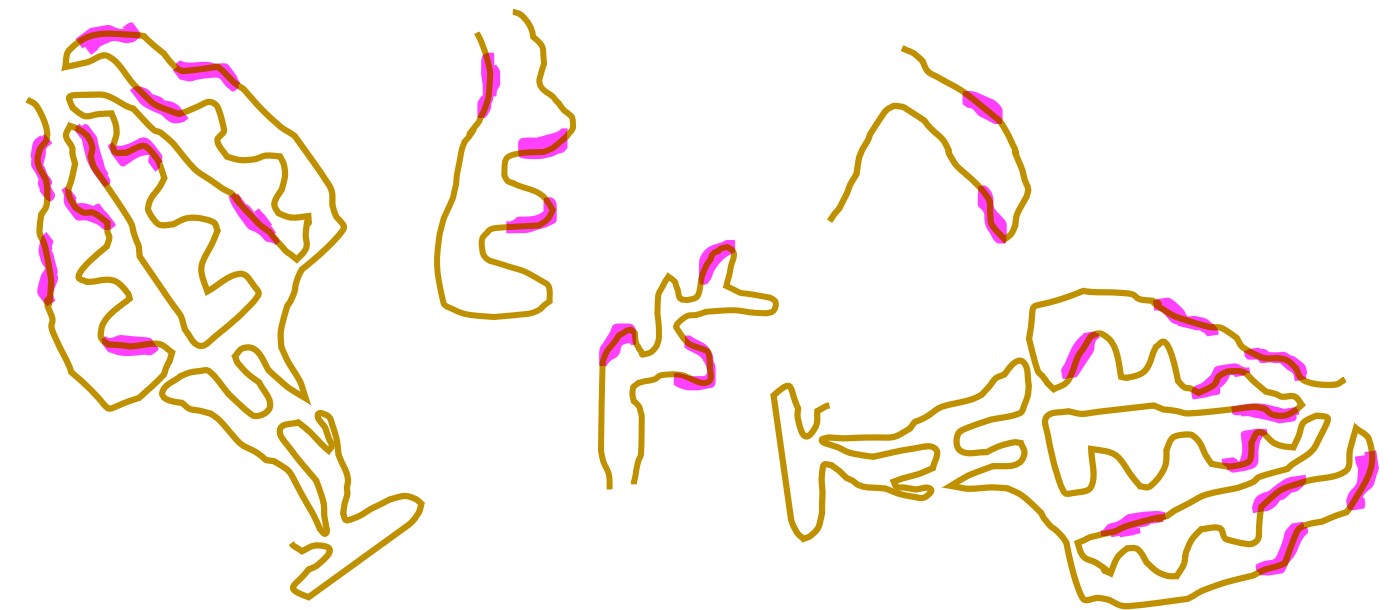
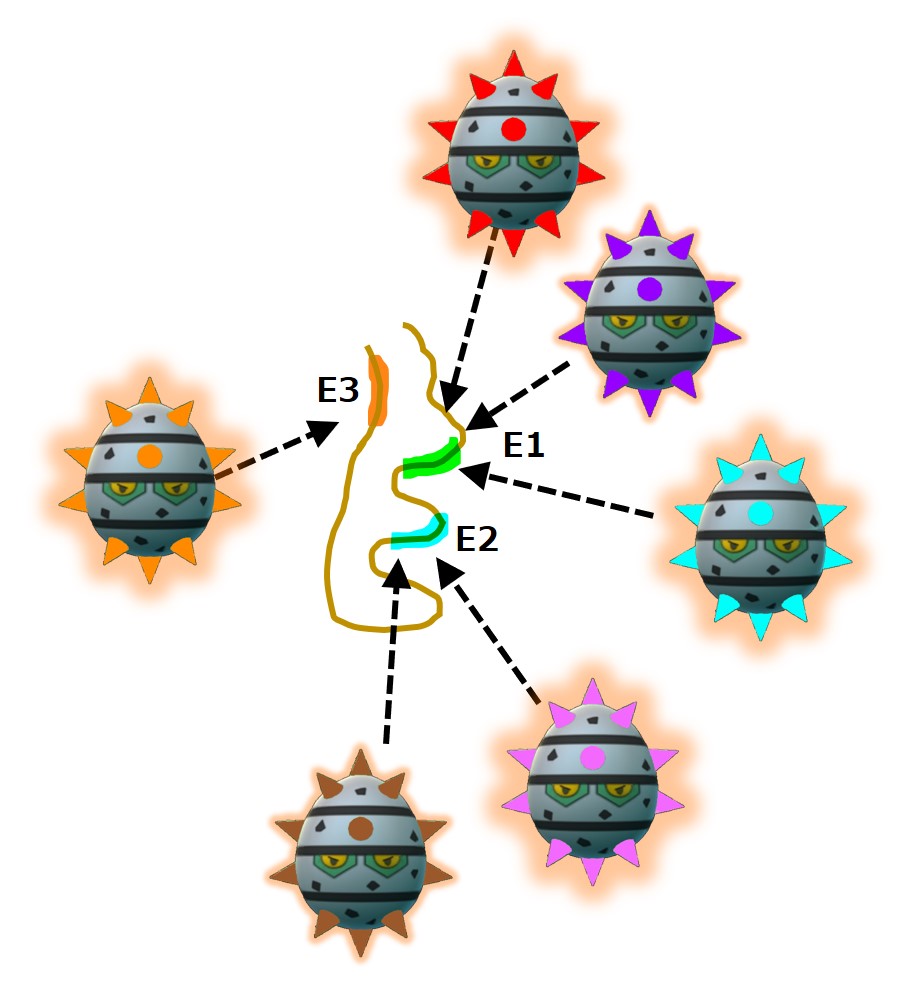
ここで、あるナイーブB細胞の抗体(B細胞受容体=IgM)が、自分の担当するエピトープにかなり似たエピトープを持つ抗原断片と結合したとします。
「3-1-2-2.抗体」の交差免疫のところで紹介したように、抗体とエピトープの接触面がピタリと合わなくても、ある程度合っていれば結合します。結合の強弱にわずかな違いが出るだけのことです。
免疫学者の概算によると、B細胞受容体のFab領域(エピトープをつかむ手の部分)のバリエーションは1000億以上あり、そのバリエーションのひとつに対してナイーブB細胞は全部で100匹くらいいるそうです。
たとえば、次図(↓)のエピトープE1に、ガッチリと完全に結合する抗体は1種類だけあり、その抗体を産生するナイーブB細胞は全身に100匹くらいいるということです。
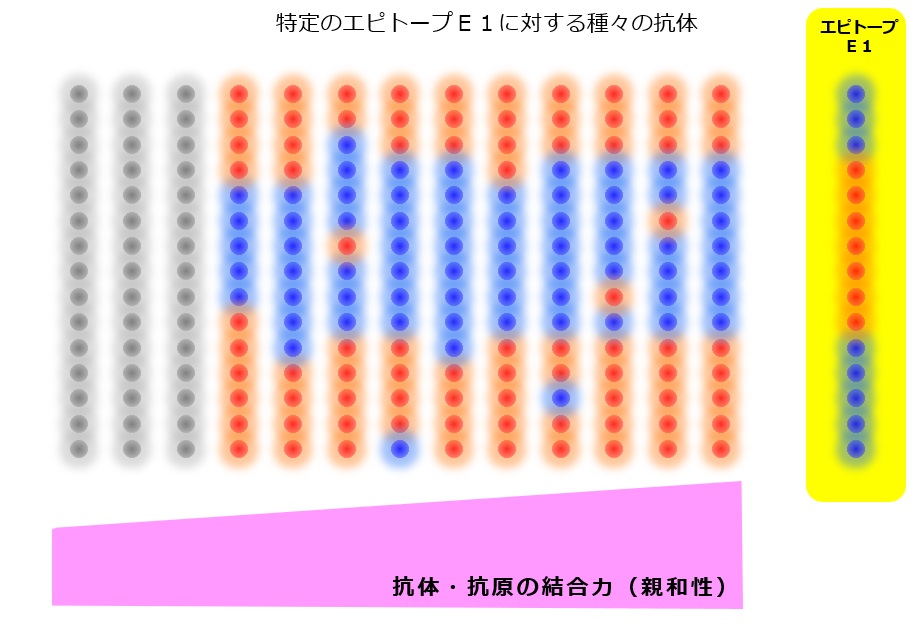
ところが、図(↑)でわかるように、エピトープE1と、完全にガッチリではないが「それなりの強さで結合する抗体」の種類は、おおよそ数千種類以上あることでしょう。
その数千種類の抗体を産生するナイーブB細胞は数十万匹いることになります。
そうすると、リンパ節に流れてきた病原体の抗原のほとんどは、数十万匹の「それなりの強さで結合する抗体」を産生するナイーブB細胞によって食べられます。
わずか100匹しかいない「完全にガッチリと結合する抗体」を産生するナイーブB細胞が病原体の抗原を食べる機会はとても少なくなります。病原体の断片を食べるナイーブB細胞全体の数千分の1を占めることになります。
ナイーブB細胞は抗体に結合した抗原断片(や病原体)を食べます。細胞の中に取り込んで消化分解します。
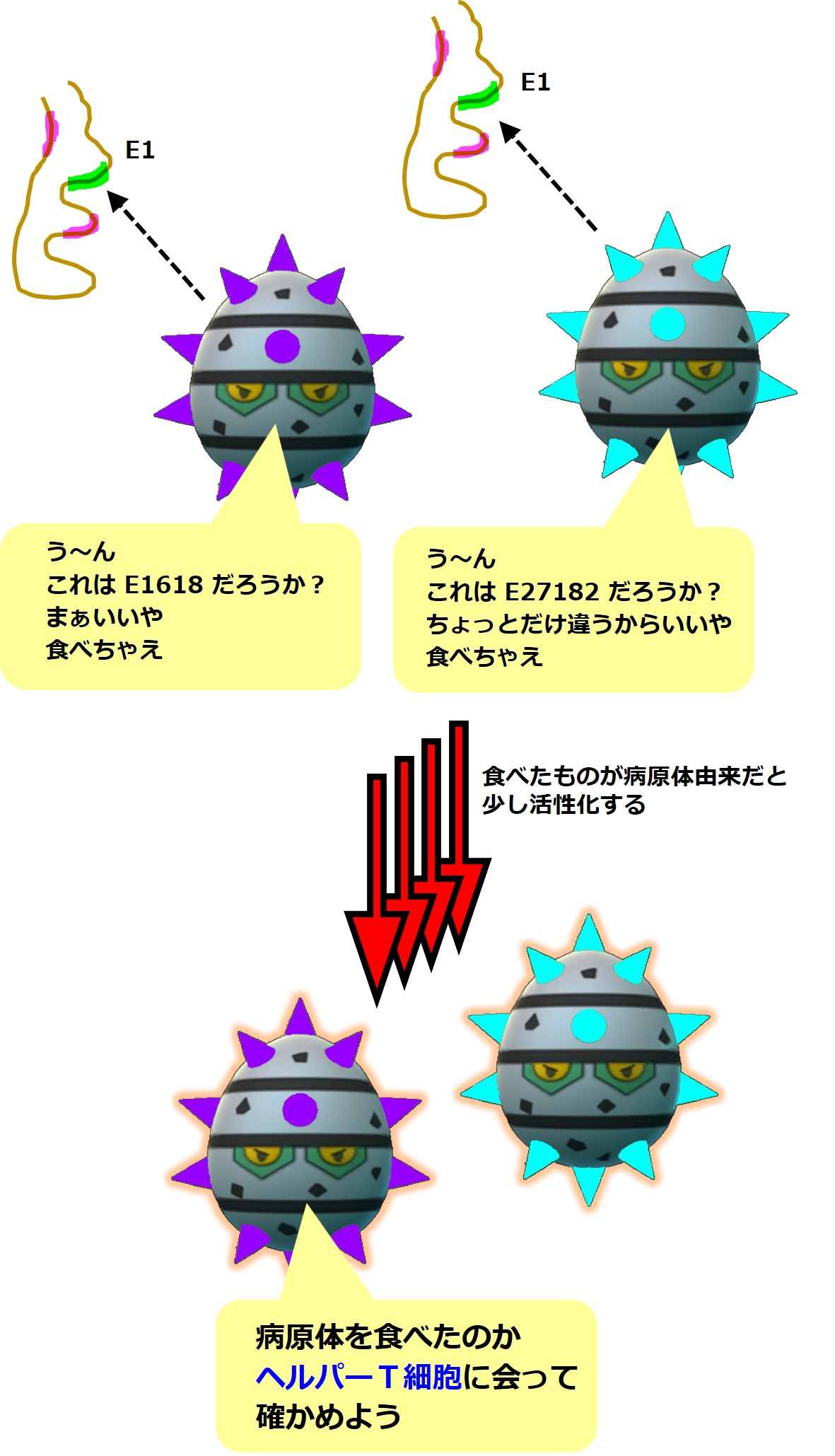
そして食べたものが病原体だと分かると、ナイーブB細胞は少し活性化します。童貞卒業が間近です。
これからいろいろな儀式を経て一人前の男になります。
その儀式は活性化ヘルパーT細胞によって執り行われます。
ナイーブB細胞は活性化ヘルパーT細胞に会うため、リンパ節内のB領域とT領域との境界へ向かいます。
リンパ節のT領域では、同じ病原体を食べた樹状細胞により、ナイーブヘルパーT細胞が活性化ヘルパーT細胞に変化しています。この活性化ヘルパーT細胞はリンパ節のT領域とB領域との境界へ向かいます。
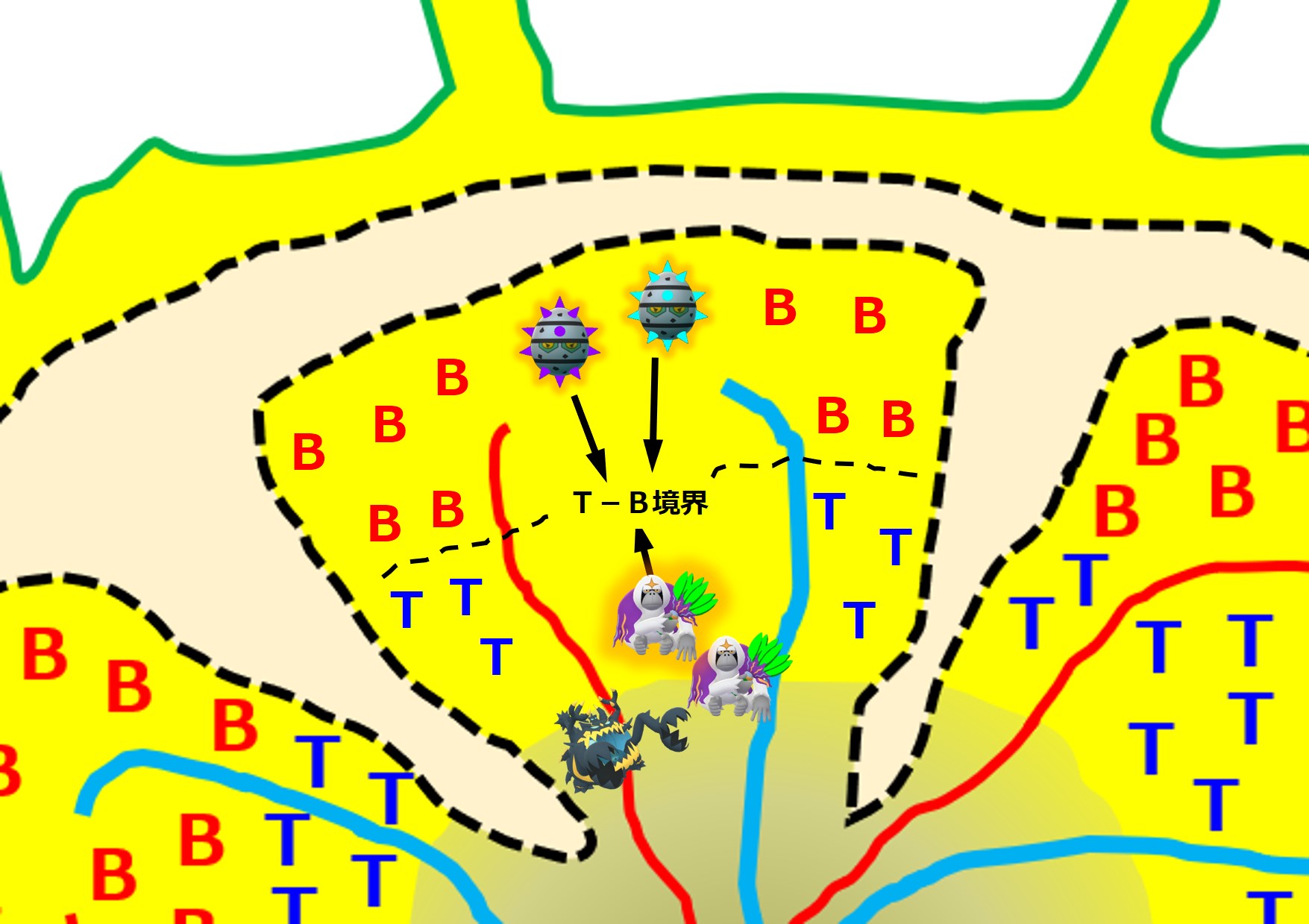
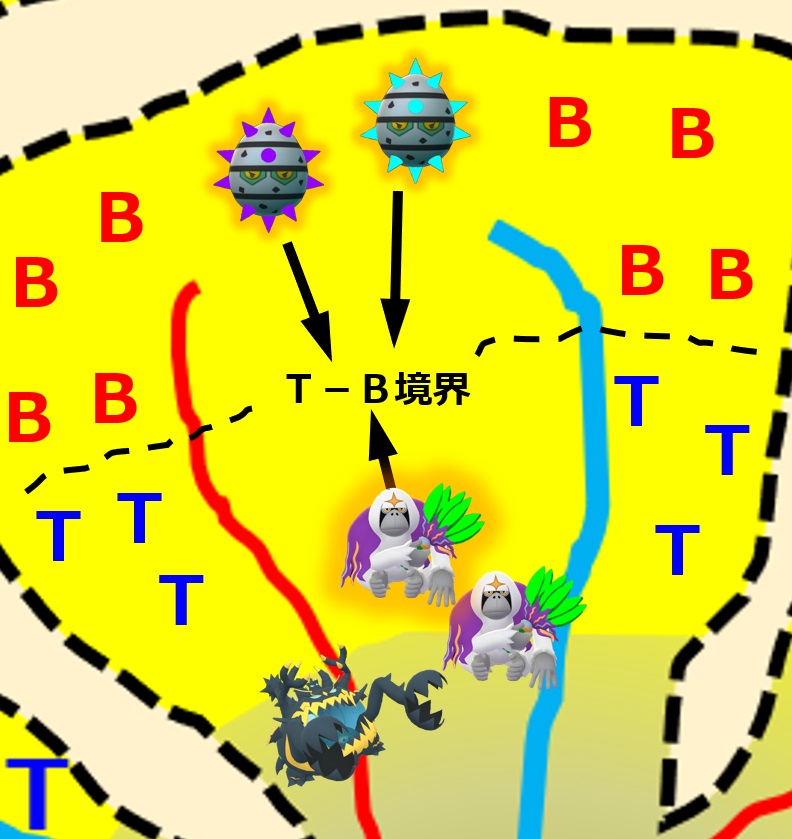
ナイーブB細胞は、バラバラに分解した抗原断片を細胞表面に提示します。「MHC クラス II + ペプチド抗原」という形で提示します。このあたりは「3-1-3.抗原提示」で説明しています。
活性化ヘルパーT細胞は「T細胞受容体」を使って、境界に集まったナイーブB細胞の提示している「MHC クラス II + ペプチド抗原」を次々とチェックしていきます。
そして、同じ種類の病原体(の断片)を食べたナイーブB細胞を見つけると、・・・いよいよ童貞卒業最後の儀式が行われます・・・ヘルパーT細胞はこのナイーブB細胞に強い刺激を与え活性化B細胞へ変えます。
さあ、こうして活性化B細胞が1匹誕生です。
実際には同じようなことがたくさん起こって、病原体が持ついろいろなエピトープに対して活性化B細胞(第1世代)が作られます。
1種類のエピトープに対して何百種類もの抗体産生B細胞が反応し、活性化B細胞になることでしょう。
特定の病原体の侵入に対して生まれた最初の活性化B細胞という意味で第1世代としておきます。
エピトープの種類が多すぎる時などは、すべてのエピトープに対して抗体ができるとは限らないようです。数の勝負なので、ナイーブB細胞の活性化は、数量の多いエピトープに対するものが優勢になるでしょう。
「同じ病原体を食べた」と活性化ヘルパーT細胞に認められて刺激を受けなければナイーブB細胞は活性化しません。
抗体を使った攻撃はとても強力なので、本当にこの抗体を産生しても大丈夫なのか、免疫系は幾重もの確認手続きを設けています。
さて、この時点で身体側はB細胞に対して何を期待しているか考えてみましょう。
B細胞は「親和性成熟」によって身体側の期待、ニーズに応えるようなことをしていますから、先にニーズを掌握しておきましょう。
そのニーズを列挙すると
結構うるさい注文ですね。こういう注文になるのは、免疫細胞が基本的には危ない連中だからです(人間でいうと人殺し、放火魔、人肉食、ゲテモノ喰い、毒コレクター、爆破マニアなど)。
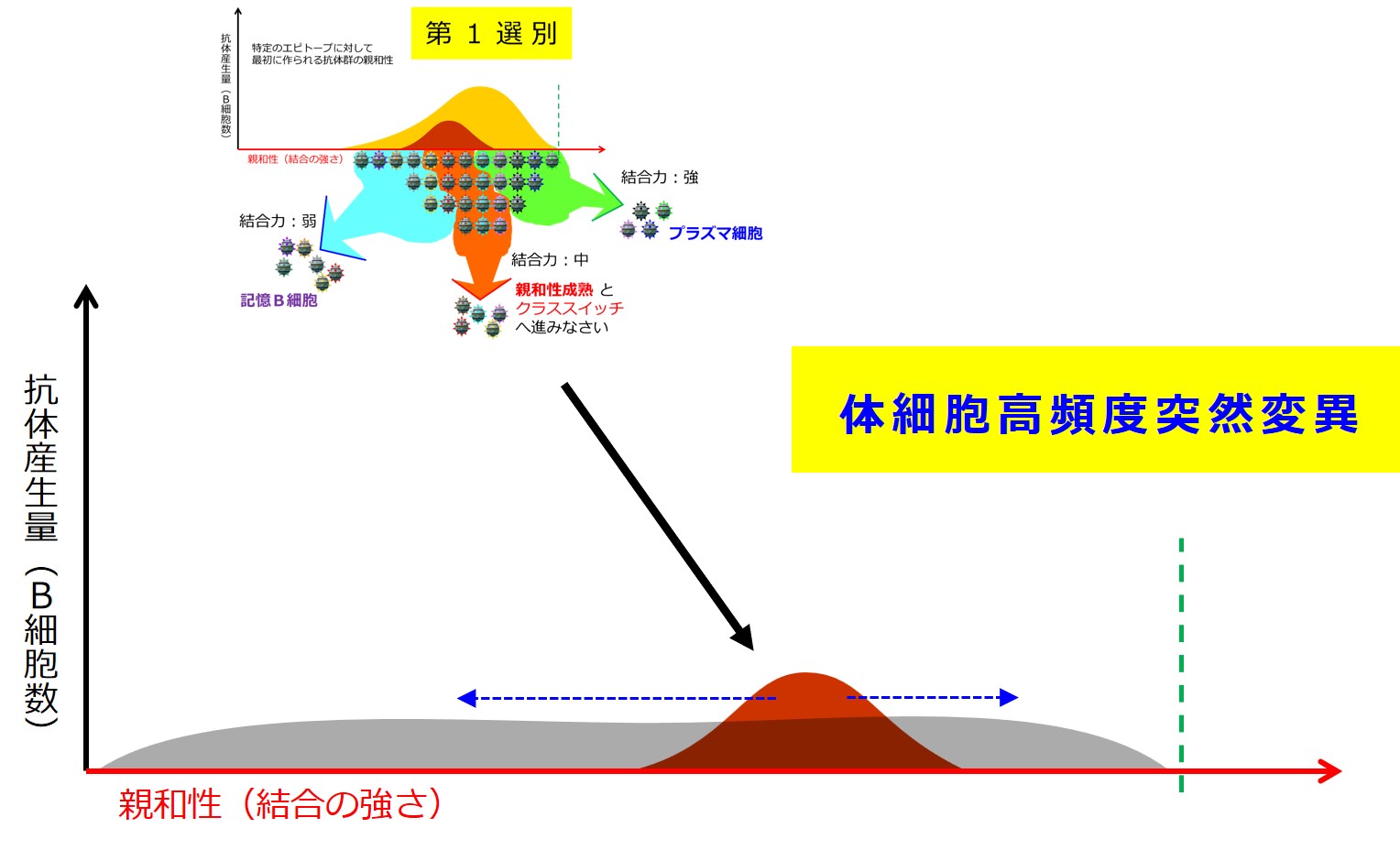
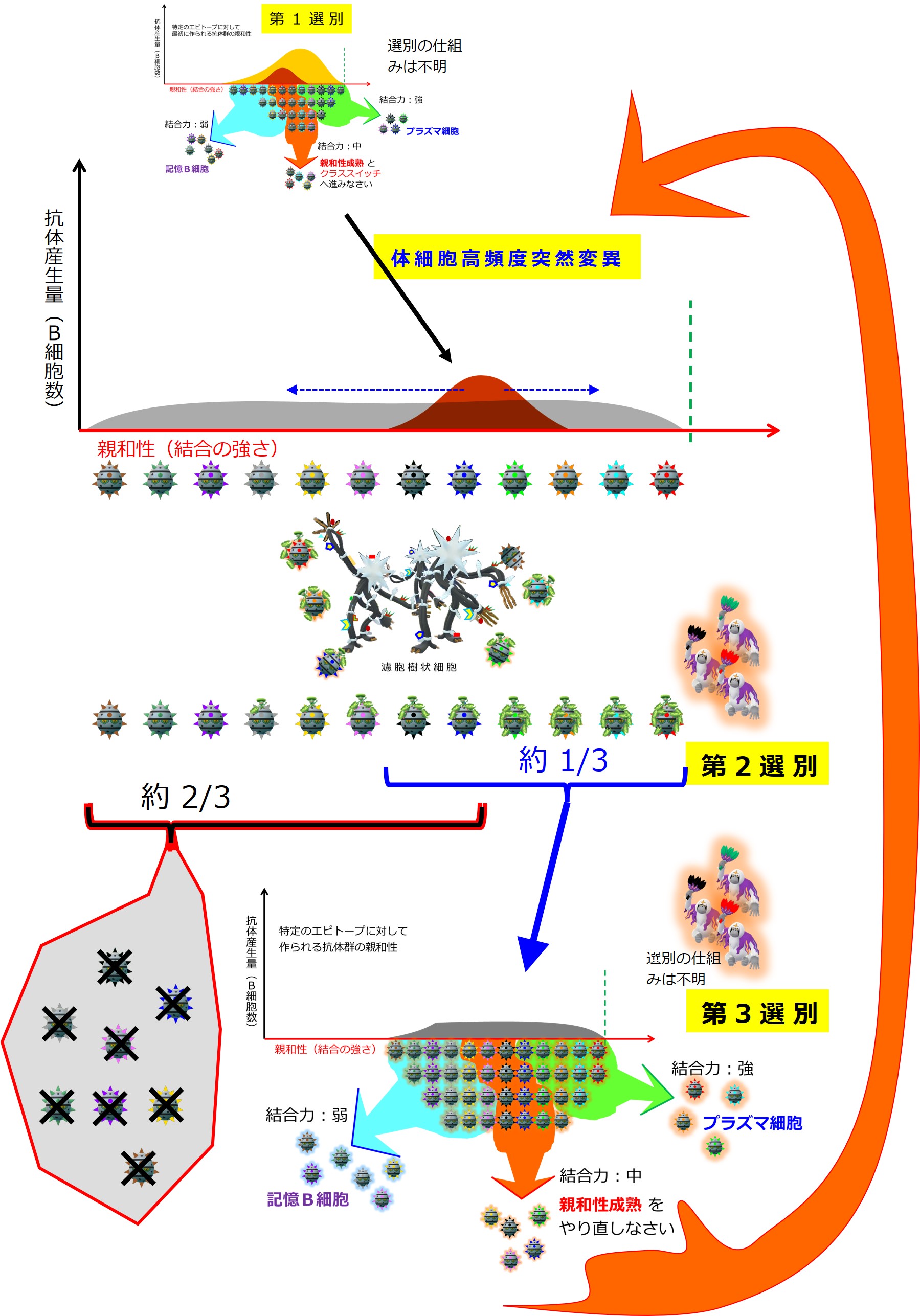
第1選別
リンパ節のT領域とB領域の境界で活性化したB細胞は、B領域(濾胞)の中心部で増殖し、大きく3つのグループに分かれていきます。
この段階の選別の仕組みはよくわかっていないようですが、B細胞受容体(細胞膜上に配置された抗体)と抗原との結合の強さで「強・中・弱」に分けられます。境界はあいまいです。
抗原との結合の強さが「強」の抗体を産生するB細胞は プラズマ細胞(分泌型抗体の産生細胞;形質細胞ともいう)に変化し、骨髄へ戻って抗体を大量に産生します。プラズマ細胞になると、もう細胞膜に抗体は発現せず、細胞外へ分泌されるようになります。増殖能力も失い、MHC II の発現もなくなります。
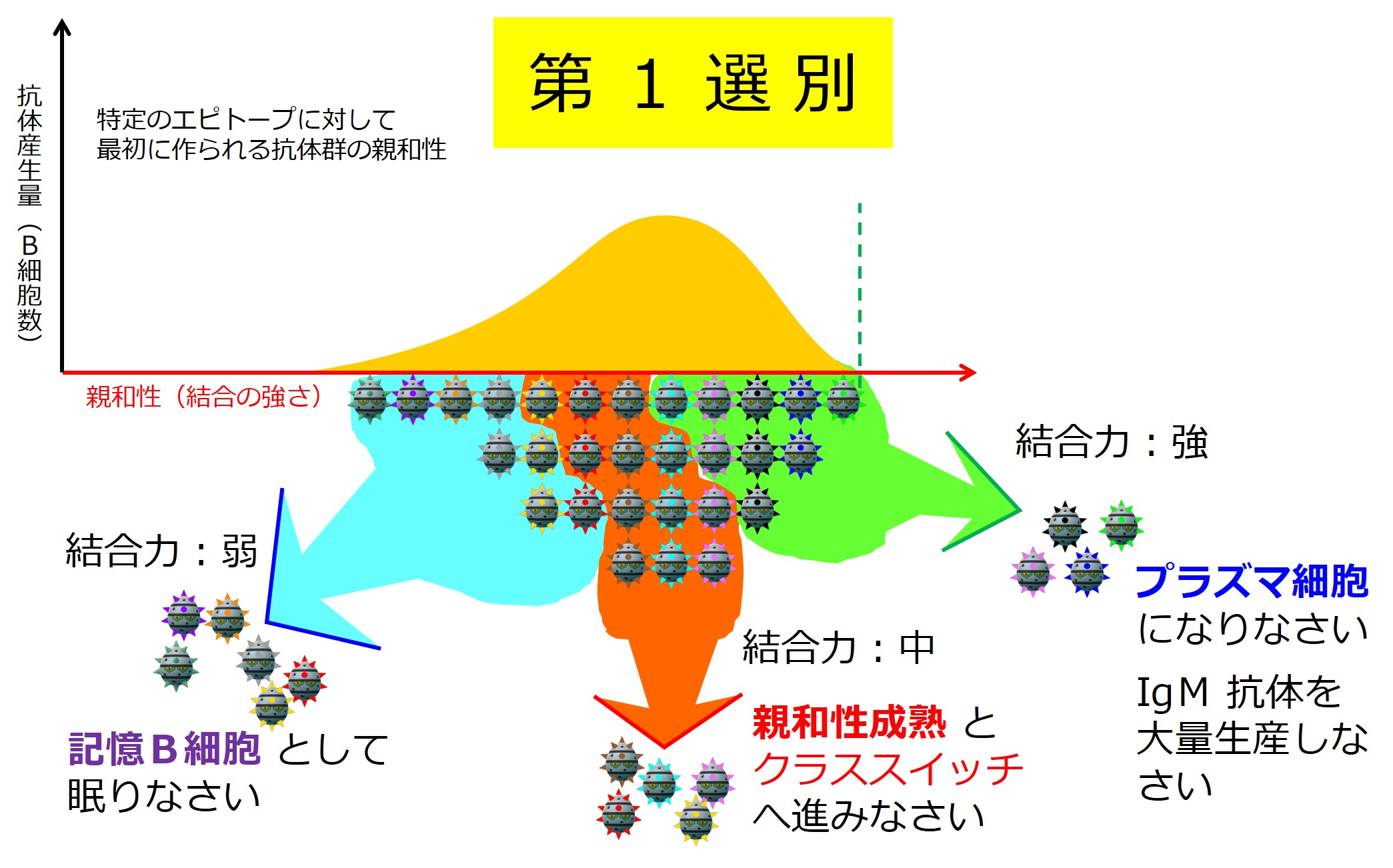
注:「第1選別」は便宜的な呼称であって免疫学用語ではありません。
| B細胞の第1選別(リンパ節) | |
| 非自己抗原との結合 | 行き先 |
| +++ | プラズマ細胞 |
| ++ | 親和性成熟 |
| + | メモリーB細胞 |
| ー | ナイーブのまま |
この抗体は「分泌型IgM抗体」と呼ばれており、5量体の形をとります。
活性化B細胞(第1世代)の中では結合力が「強」ですが、親和性成熟を経た活性化B細胞(第2世代以降)が産生する IgG より結合力は弱いです。
ニーズ1に応えるために、取り急ぎ作られる抗体です。
血液中に IgM抗体 が出現するのは、自然免疫(食細胞)と病原体との激しい戦いが始まってから4~5日後のことです(小競り合い程度だと獲得免疫系は動き始めません)。
この IgM産生プラズマ細胞 の寿命は2~3日と短く、次の親和性成熟とクラススイッチを経て(戦いの開始から10日後以降に)出現する IgG産生プラズマ細胞(長寿命:数か月~数十年)と置き換わっていくことになります。
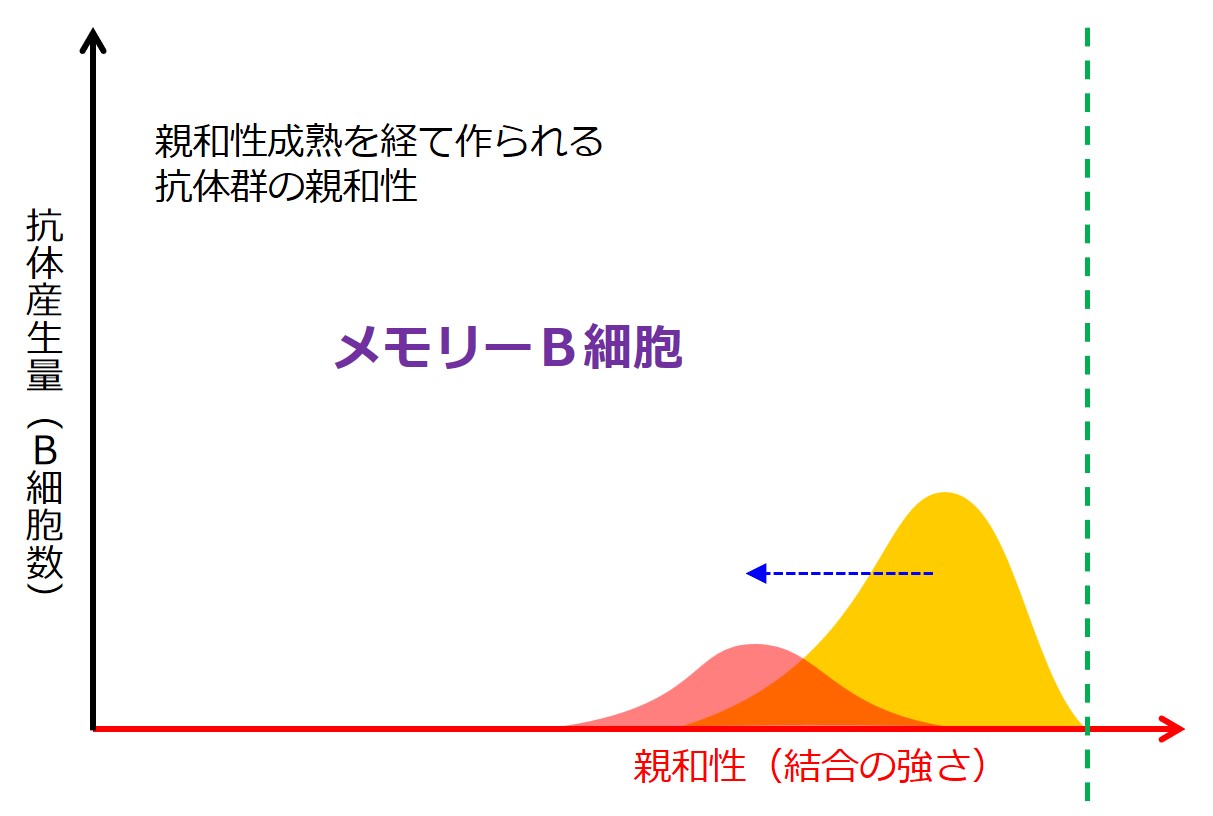
第1選別において抗原との結合力が「弱」の抗体を産生するB細胞はメモリー細胞(記憶B細胞)になります。[編集メモ:これがIgG産生B細胞へクラススイッチされているのか、それともIgM産生B細胞のままなのかは不明:要確認]
次が「親和性成熟」の入り口となります。なぜわざわざ入り口を明示しているのかと言いますと、親和性成熟コースで、とても良い成績を修めたB細胞は抗体産生へ進み(おめでとう!)、とても出来の悪かったB細胞は殺されますが(ありがとう、ご苦労様でした、もういいよ!)、ほどほどの成績のB細胞は「親和性成熟コースのやり直し」を命ぜられます(まだチャンスはあるよ、頑張ろう!)。この入口に戻ってくることになるので、入口を明示しています。
第1選別(あるいは第3選別)においてエピトープとの結合力が「中」のB細胞は、すこし大げさに言うと、濾胞の中心部の光の届かない暗領域(ダークサイド)へ移動し、突然変異で化けながら大増殖して 胚中心 と呼ばれる構造を形成します。ここでは普通の突然変異の百万倍もの高確率で変異が起こるそうです(体細胞高頻度突然変異 somatic hypermutation と呼ばれるようです)。
この滅茶苦茶な突然変異は、抗体のFab領域をコードしている遺伝子に起こるようです。突然変異によって増殖したB細胞の作る抗体の結合能力は強弱様々なものに変化します。
もともとのエピトープにかなり強く結合する抗体を作れるようになったB細胞、むしろ弱く結合する抗体を作るようになったB細胞、あるいはまったく結合できない抗体を作るようになったB細胞などが現れます。
また、この胚中心が形成される前に「IgM産生 → IgG産生」へと クラススイッチ も起こっているようです。いつ、どのようにして起こるのか詳細はまだ不明のようです。
骨髄の学校で「自己抗原とガッチリ結合する自己抗体産生B細胞」は殺されたのですが、この体細胞高頻度突然変異は、いわばリセットですから、また「自己抗体」を産生するB細胞が生まれる可能性もあります。
変異B細胞の選別は、胚中心の明領域と呼ばれる明るい場所で行われます。
胚中心の明領域には 活性化ヘルパーT細胞 と 濾胞樹状細胞(ろほうじゅじょうさいぼう:FDC;Follicular dendritic cells)と呼ばれる細胞がいます。
濾胞樹状細胞 FDC は、自然免疫に属する樹状細胞 DC とは名前が似ているだけで、別種の細胞です。
明領域の活性化ヘルパーT細胞と濾胞樹状細胞は、変異B細胞を選別する試験官のような役割を担っています。
この濾胞樹状細胞 FDC は、リンパ節に流れてくる抗原のうち 抗体や補体の結合した抗原 を拾って自分の体に貼り出す習性を持っています。抗原のショーウインドウ細胞 とも呼ばれているようです。抗原を食べて分解しているわけではありません。生の抗原をそのまま貼りつけているのです。
病原体が細菌だと直接補体が結合することもありますが、ウイルスの場合は抗体の結合が必要でしょう。そうすると、先に分泌される IgM の働きがあって親和性成熟を始めることができることになります。
壊れた体細胞の断片など、自己抗原が流れてきても、自己抗体をたくさん持っていないときは抗体が結合していないので、自己抗原が濾胞樹状細胞に拾われることはありません。
暗領域から明領域に移ったB細胞の抗体(B細胞受容体:IgG)は、濾胞樹状細胞 FDC の展示している抗原に、結合するものもあれば、結合しないものもあります。結合の強さも様々です。
濾胞樹状細胞 FDC の展示抗原と強く結合したB細胞は、展示されていた抗原を食べます。B細胞は食べた抗原を消化分解し、「MHC クラス II + ペプチド抗原」を細胞表面に出します。抗原をたくさん食べたB細胞は「MHC クラス II + ペプチド抗原」をたくさん提示します。
濾胞樹状細胞 FDC の展示抗原と弱く結合したB細胞も、同じように反応しますが、抗原をたくさんは食べることはできないので「MHC クラス II + ペプチド抗原」を少しだけ出します。
濾胞樹状細胞 FDC の展示抗原とまったく結合できなかったB細胞は、抗原をいっさい食べないので「MHC クラス II + ペプチド抗原」をまったく提示しません。
結合の強さによってB細胞膜上に出てくる「MHC クラス II + ペプチド抗原」の数が異なってきます。
明領域にいる活性化ヘルパーT細胞がB細胞の選別をおこないます。
第2選別
B細胞膜上に出ている「MHC クラス II + ペプチド抗原」の数が多い上位 1/3 は、第3選別へ進みますが、数の少ない下位 2/3 は殺されます。アポトーシス(細胞の自殺)のかたちで死滅しますが、どの細胞の指示で事が進むのか機序は不明です。
キラーT細胞は居ないようなので、明領域にいるヘルパーT細胞がB細胞膜上に出てくる「MHC クラス II + ペプチド抗原」の密度に応じてB細胞の選別をおこなっている可能性があります・・・が不明です。
下位 2/3 の排除はニーズ4(自己抗体を作らない)に対応するためでしょう。
| B細胞の第2選別(リンパ節) | ||
| 割合 | ろ胞樹状細胞の 展示抗原との結合 |
行き先 |
| 1/3 | +++ ~ ++ | 第3選別へ |
| 2/3 | + ~ ー | 死滅 |
第3選別
第2選別で生き残った「MHC クラス II + ペプチド抗原」の数が多い上位 1/3 は、(第1選別と同じように)抗原との結合の強さで「強・中・弱」に分けられ、「強」のB細胞はプラズマ細胞に変化し、骨髄へ戻って抗体を産生します。結合力が「弱」のB細胞はメモリー細胞(記憶B細胞)になります。結合力が「中」のB細胞は、濾胞の暗領域(ダークサイド)へ戻ります。「親和性成熟」の入り口に戻り、突然変異からやり直します。
この段階の活性化B細胞、親和性成熟を1度くぐったB細胞が第2世代ということになります。2度くぐると第3世代、3度くぐると第4世代・・・体内の病原体が完全に駆逐されると親和性成熟はできなくなります。
| B細胞の第3選別(リンパ節) | ||
| 割合 | 非自己抗原との結合 | 行き先 |
| 1/3 | +++ | プラズマ細胞 |
| 1/3 | ++ | 親和性成熟やり直し |
| 1/3 | + | メモリーB細胞 |
第1選別と似たようなことが行われます。
詳細な仕組みは不明です。
「第1選別~第3選別」は、便宜的呼称であって
学術的な呼び方ではありません。
第2選別と第3選別の区分は(理解を容易にするための)便宜的なものであり、実際には同じ場所で同時におこなわれている選別かもしれません。
第2選別と第3選別を掛け合わせると次表のようになります。
| 親和性成熟におけるB細胞の選別(リンパ節) | ||
| 割合 | 病原体との親和性 | 行き先 |
| 1/9 | 高い | プラズマ細胞となり抗体を多量産生する |
| 1/9 | 中くらい | 親和性成熟へ進み性能向上をめざす(やり直し) |
| 1/9 | 低い | 記憶B細胞となり眠りにつく |
| 6/9 | 無い | 死ぬ |
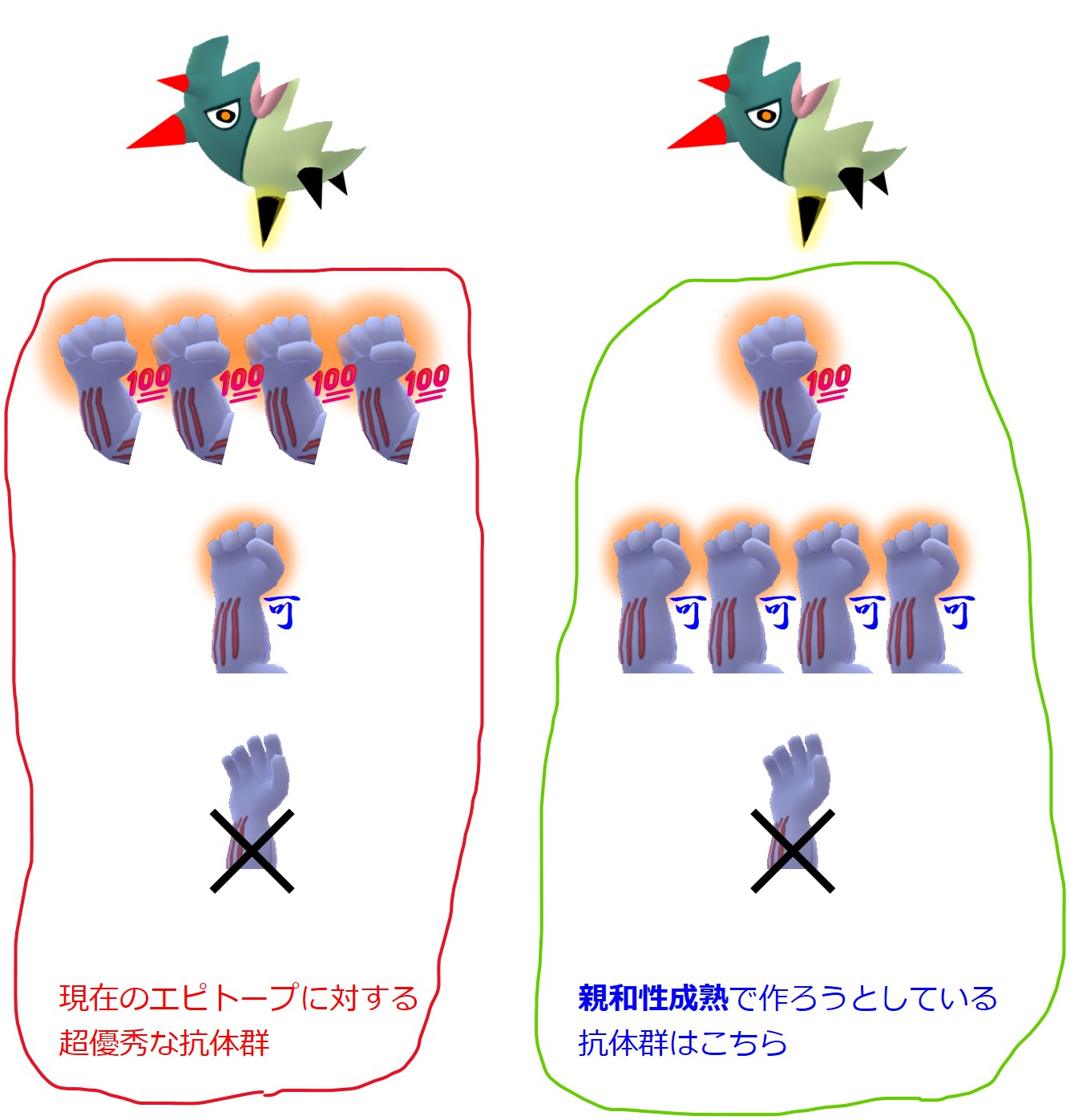
この親和性成熟を経て、病原体のエピトープとガッチリと強く結合する、つまり、間違っても自己抗原とは結合しない抗体の産生が可能になります。
しかし、この親和性成熟の仕組みを見てわかることですが、親和性成熟を繰り返すことで「病原体の抗原(エピトープ)と完璧にガッチリと結合する百点満点の抗体」を産生するB細胞を作ろうとか、殖やそうとかはしていません。
親和性成熟の繰り返しで、あるエピトープにかなり強く結合するけれども多様な手をもつ抗体産生細胞群を作ろうとしています。
この多様な手は、エピトープのちょっとした変異には十分に対応できるでしょう。
免疫にとって未経験の新しい病原体に初めて感染したとき、発症の4~6日後、血液中に出現する抗体は IgM です。そのころリンパ節では親和性成熟とクラススイッチが行われており、数日後には血液中に IgG が出現します。
「3-1-2-3.抗体の種類」でみたように、IgG の結合した抗原に対して免疫系は多彩で強力な方法で攻撃を仕掛けます。
同種の病原体による再感染時には、IgG産生のメモリーB細胞が素早く増殖するので、IgM は産生されないか、あるいは IgM と IgG がほぼ同時に血液中に出現するようです。メモリーB細胞は活性化に必要な手続きが簡略化されているようです。

また、消化管等のリンパ節では「IgM産生 → IgA産生」へのクラススイッチがおこり、IgA産生型のプラズマ細胞は粘膜下へ移動し、そこで分泌型 IgA(二量体)の産生・分泌をおこないます。
消化管等の粘膜細胞は、この IgA を取り込み消化管等の内腔側に運びます。
なお、健康な成人は毎日約3グラムの抗体を産生しているようですが、その 2/3 は IgA だとのことです。
IgAは食物等と共に流れ去りますから、たくさん作らざるを得ませんね。
免疫系が身体のニーズを満たしているか再確認してみましょう。
| 要求1 | ともかく、性能は問わないから一刻も早く抗体を産生して病原体の増殖を抑えてほしい。とにかく抗体の姿を出してくれ。 | |
| 性能1 |
|
|
| 要求2 | 病原体を強烈に攻撃する免疫応答を呼び込む抗体(つまり IgG)が間違って自己細胞を攻撃すると超危険なので、そういう抗体は確実に、ピンポイントで病原体のエピトープとだけ結合するようにしてほしい。 | |
| 性能2 |
|
|
| 要求3 | 病原体はまた来る、変異して来るだろう。次の感染予防に使う抗体は結合するエピトープに幅を持たせ、少しの変異には柔軟に対応できるようにしてほしい。 | |
| 性能3 |
|
|
| 要求4 | かといって、どんな大きな変異にも対応しようとすると自己抗原に結合する危険もあるから、それは絶体にやめてほしい。エピトープの幅を拡げすぎないでほしい。 | |
| 性能4 |
|
|